ウミウシのダンスとSNS

ウミウシの交尾について話したい。
地中海の浅瀬に、エリシア・ティミダという体長2センチほどのウミウシがいる。背中に緑の模様、頭部に小さな赤い斑点。藻類を食べ、その葉緑体を体内に保持して光合成する。
ウミウシのほぼすべての種は、同時的雌雄同体だ。一個体がオスとメス両方の生殖器官を同時に持ち、卵も精子も作る。だから交尾は、お互いがオスとしてもメスとしても振る舞いながら、精子を交換しあう作業になる。
エリシア・ティミダが交尾をするときの流れを説明しよう。
二匹は出会うと頭をぶつけ合う。それから体を反対方向に向けて並ぶ。ペニスを伸ばし、相手の背中に注射する。針もないのにどうやって皮膚を貫通するのかは、「未解明」とされている[1]。短いと数秒、長いと30分以上の注射を終えると、二匹は同期して反時計回りに円を描く。ただひたすら一緒に回る。回り終わるとまた注射する。これを繰り返す。回数は、ペアによって平均13回、最大34回に達する。最後にようやく、お互いの生殖孔同士で精子を交換し、別々の方向に去っていく。全行程は平均で43分。
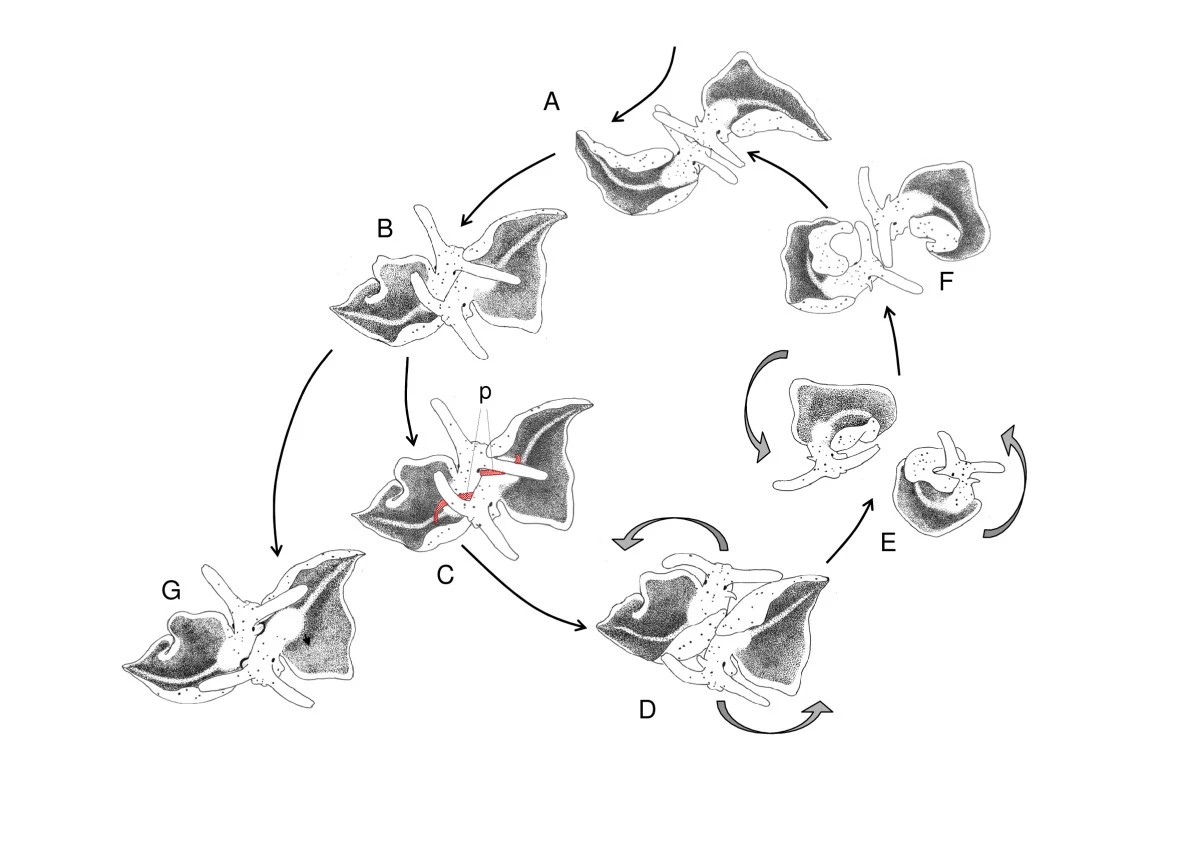
Mating sequence of E. timida. A Head-to-head contact B Copulation position C Hypodermic transfer, showing extended penises (p, marked red) D Start of circling E Circling F End of circling G Sperm transfer into female aperture. Drawings by V. Schmitt from video recordings.
ドイツのミュンスター大学、ヴァレリー・シュミットらの研究グループによれば、注射の回数も時間も、ペア同士ではほぼ均等であるという。片方が長く注射すれば、もう片方も長く注射する。最後の精子交換は、ほぼ同時に起きる。研究では、同期しなかった2例が報告されている。このケースにおいては、片方が応答するまで相手が約30分も円運動を続けて待ったという[1]。
先の研究では、この現象を「均衡互酬性(balanced reciprocity)」と呼び、その背後に「条件付き互酬性(conditional reciprocity)」のメカニズムがあると論じている。お互いの行動を見ながら、どこまで投資するかを微調整する仕組み。一方的に搾取されないように、降りる権利を保持しながら、ゆっくり時間をかけて関係性を構築していく。
四つの様式
ウミウシたちの交尾様式は、他にもある。

イラストは雰囲気で作成しており、生物学的にはかなり不正確
インド・西太平洋に広く分布し、日本では本州中部以南から琉球列島で見られるチリメンウミウシは、まったく別の方法を進化させた。大阪市立大学の研究グループによると、このウミウシは交尾のたびにペニスを切り捨てるという[2]。体内には3回分の長さのペニスが、らせん状に圧縮されて格納されている。切り捨てたあと、次の部分が24時間ほどで使えるようになる。
切り離されたペニスを調べると、表面には逆向きの棘が生えており、そこに精子の塊が引っかかっている。交尾相手の体内にすでに貯蔵されている、ライバルの精子を棘によって掻き出していると考えられている。
オーストラリア北東部のグレートバリアリーフ周辺に生息するウミウシ(暫定的に Siphopteron sp. 1 と呼ばれている)は、よりアグレッシブな交尾の様式を持っている。このウミウシは、ペニスとは別の、針のような器官を相手の額に深く挿入し、前立腺由来の分泌物を中枢神経系のすぐ近くに注入する[4]。研究グループはこれを「頭部外傷性分泌物注入(cephalo-traumatic secretion transfer)と命名している。注入される物質が相手にどう作用するかはまだ実証されていないが、同じ雌雄同体の陸産巻貝の研究では、注入される分泌物が受容者の生理に作用することが知られている。
広義のウミウシに含まれるアメフラシは、ペアではなく集団で交尾する。3匹以上が連鎖し、「デイジー・チェーン」と呼ばれる構造を作る。先頭の個体はメス役のみ、最後尾はオス役のみ、中間の個体はオスおよびメスとして機能する。アメフラシの場合、ペニスは頭部の右側、メスの生殖孔は外套腔の奥にある。この配置のせいで、ペアでは同時に相互交尾ができない。そこでチェーン状になる[5]。
対立の構造
アメリカの生物学者ジャネット・レナードは、「雌雄同体のジレンマ(Hermaphrodite's Dilemma)」と呼ばれる対立を定義した[6]。同時的雌雄同体は、両方の性機能を持つ。だが両者ともオス役(精子提供側)を好み、メス役(受容側)を避けたがる。卵を作るコストが高い。種によっては、外傷性交尾で身体を傷つけられたり、受け取った精子の管理にコストがかかったりする。受け身の側は搾取されやすい。レナードはこのジレンマを整式化した。同時的雌雄同体の交尾行動はすべて、この対立への解として理解できる。
エリシア・ティミダの条件付き互酬性は、長い時間をかけて少しずつ投資し、相手も同じだけ投資することを確認しながら進める解。チリメンウミウシの使い捨てペニスは、過去の他者の精子を物理的に消去することで自分の遺伝的成功を確保する解。Siphopteron sp. 1 の頭部への注射は、相手の生理を化学的に操作する解。アメフラシの連鎖交尾は、生まれ持った制約のもとで複数の個体が協働し、役割を順送りにする解。
このうち、エリシア・ティミダの交尾の様式は、関係を壊さない仕組みを行動のなかに組み込んでいる点で特殊に見える。
たとえば、SNSで誰かの発言に反論する。相手は強く言い返してくる。こちらはもっと強く出る。相手も強く出る。最初の発言から離れて、関係そのものが燃える。応答が応答を呼び、累積してエスカレートする関係を、人類学者グレゴリー・ベイトソンは分裂生成(schismogenesis)と呼んだ[7]。これに対して、相手の出方に合わせて自分の出方を引いたり戻したりする——応答が暴走しないように、互いに調整し続ける関係が互酬性(reciprocity)だ。エリシア・ティミダの注射と円運動の交替は、後者にあたる。両者が同じだけ投資する。片方が暴走すれば、もう片方は応答を止める。それで関係が壊れない。
もちろん正解はない。それぞれの種が直面する密度・遭遇率・捕食圧・寿命という条件のもとで、別々の解が生まれた。「あるべき交尾」はない。
ウミウシのような設計
ウミウシは交尾しているのであって、会話をしているわけではない。それでも「ウミウシの交尾のようなSNS」を考えてみたい。
エリシア・ティミダ型:沈黙の同期と、意味に回収されないコレオグラフ
エリシア・ティミダの交尾の核心は、注射と注射のあいだに起きる円運動だ。二匹は反時計回りに回る。情報を交換しない。発言もメッセージも何もない。ただ一緒に回るだけ。これが終わると、また注射する。
関係の構築のなかに、意味の応酬(体液の交換)が止まる時間が埋め込まれている。沈黙とコレオグラフが、関係を成立させる構造的な要素になっている。
いまのSNSは、意味の応酬が止まることを「離脱」「無関心」「関係の終わり」として読む。既読がついて返事がないことは、関係の停滞のサインだ。沈黙はネガティブな状態として扱われている。
通りで他人とすれ違うとき、人は会話をするわけではない。それでも互いに距離を取り、視線を外し、相手が動けば自分も少し動く。意味はないが、調整はある。カナダの社会学者アーヴィング・ゴッフマンはこうした「会話のない調整」を 「焦点の定まらない相互作用」と呼び、会話のように共通の話題に向けて協調する状態(焦点の定まった相互作用)と区別した[8]。エリシア・ティミダの円運動は、焦点の定まらない相互作用にほぼそのまま当てはまる。
いまのSNSが設計の対象にしているものの多くは、「焦点の定まった相互作用」だ。投稿、いいね、返信、引用、批判——どれも、ある発言を中心に別の誰かが応じる構造で動いている。
エリシア・ティミダ型のSNSを考えてみよう。メッセージのやりとりに、構造として「同期した沈黙」や「身振り」のフェーズが組み込まれる。意味の応酬の合間に、一定時間、両者が一緒に何もしないことが画面上で可視化される。お互いがそこに「いる」ことだけが共有される。それが終わると、また往復が始まる。
円運動は、相手の動きに自分の動きを合わせる。エリシア・ティミダの研究には、印象的な観察がある。最終局面で一方しか精子を渡さなかった2例において、円運動が約30分にわたって続いた。通常ならば、精子交換のあとすぐに分離する。関係が崩れかけたときに「沈黙の動機」が異常に長くなる、と考えることもできる。
設計に置き換えれば、応答が滞ることは「離脱」ではなく「関係の異常を示すサイン」と解釈することになる。意味に回収されない時間の長短が、関係のバランスを測る指標になる。
もう一つ、エリシア・ティミダ的な設計要素を考えるならば、「背中」だ。
注射は背中側に行われる。生殖孔ではない場所に、何度も、交互に繰り返され、それから本格的な精液の交換が起きる。これを設計に置き換えれば、メインのメッセージ機能とは別に、ささやかな印を残せる場所に「身体」を持たせるという思想になるだろう。ホームでもプロフィールでもDMでもない、相手の「背中」に当たる領域を用意する。そこに何度も触れあうような設計。何が伝わっているかは、わからなくていい。
チリメンウミウシ型:自分の一部を残して離脱する
チリメンウミウシの使い捨てペニスを設計の参照として読むなら、自分の身体の一部を切り離して、相手の中に置いてくるという構造になる。
ペニスは切り離されたあと、棘によって相手の内部に固定される。持ち主はそこから去る。だが自分の一部は、相手の中に残り続ける。痕跡が残ることと、自分から切り離すことが、同時に起きている。
いまのSNSの「離脱」には、二つの極しかない。すべてが残るか(発言は永続する)、すべてが消えるか(削除する/される)。「自分の一部を相手のなかに置いて、本体は去る」という第三の様式はまだ普及していない。
チリメンウミウシ型の設計は、この第三の様式を実装する。会話を終えるとき、自分の発言の一部を「相手のなかに残るが、自分の統治下にはないもの」として、能動的に切り離す。発言は残る。だがそれはもう自分のものではない。修正もできない。削除もできない。返信を求められても応じない。発言は相手の所有物になり、自分は立ち去る。
この設計が解くのは、所有をめぐる対立だ。発信したユーザーは所有権を譲渡することで、痕跡を残したまま離脱できる。内容は相手のなかに残り続けるが、最初の発信者の介入から自由になる。
アメフラシ型:位置が役割を決める
アメフラシのデイジー・チェーンでは、チェーンにおける位置が、その個体の性的役割を決める。先頭の個体は受信のみ。末尾は発信のみ。中間にいる個体だけが、両方の役割を同時に担う。
アメフラシの場合、この役割分担はペニスと生殖孔の解剖学的な配置から生み出された帰結だ。SNSの設計に借りるなら、解剖学に代わる別の論理で位置と役割の対応を作る必要がある。それは時間か、他の参加者との関係か、コミュニティ内の経歴か。
いまのSNSは、ひとりの参加者が常に発信者でもあり受信者でもある、という前提で設計されている。投稿もできる、見ることもできる、両方が同時に開かれている。
アメフラシ型の設計は、これを否定する。ある会話のなかで、参加者は「いまの自分の位置」によって、発信のみ、受信のみ、両方、のいずれかになる。位置は固定されない。時間とともに移動する。先頭にいた人が中間に移り、最後尾になり、また先頭に戻る。
たとえば、こんな実装が考えられる。会話空間に入ったとき、参加者は「いまは聞くだけ」「いまは話すだけ」「いまは両方」のいずれかの位置を割り当てられる。位置は他の参加者との関係で決まる。最も新しく入った人は受信のみ、最も古くから話している人は発信のみ、中間にいる人だけが両方できる。
この設計が解くのは、常に両方できることで生まれる過剰さだ。
発信と受信が同時に開かれていることは、豊かなユーザー体験をもたらすように思える。しかし実際には、多くのユーザーが「常時反応を引き出されている」状態で、休む「位置」がない。
借用しにくいもの
四つの様式のうち、Siphopteron sp. 1 の頭部注射だけは、設計の発想として借りることが難しい。相手の認知に直接介入し、判断を書き換えるという構造は、振る舞いを変える操作という点で、現代のアルゴリズム配信や心理操作型の広告と問題系が重なる。生物の世界でのアグレッシブな解が、人間社会では肯定的ではない形ですでに一部実装されている。ウミウシから取り出せる発想のすべてが、素晴らしいものというわけではない。
意味の手前で
できることならば、ウミウシになりたい、と思うことがある。意味に回収される以前の場所で、等価交換のダンスを踊ってみたい。ナイーブな妄想だろうか。
でも、サービスやプロダクトの設計レベルならば、それができるかもしれない。
私たちが使っているコミュニケーションツールは、別の様式であり得た。
ニュースレター、登録は無料です。よろしければ。
脚注
[1] Schmitt, V., Anthes, N., & Michiels, N. K. (2007). "Mating behaviour in the sea slug Elysia timida (Opisthobranchia, Sacoglossa): hypodermic injection, sperm transfer and balanced reciprocity." Frontiers in Zoology, 4:17. https://doi.org/10.1186/1742-9994-4-17
[2] Sekizawa, A., Seki, S., Tokuzato, M., Shiga, S., & Nakashima, Y. (2013). "Disposable penis and its replenishment in a simultaneous hermaphrodite." Biology Letters, 9(2):20121150.
[3] Sekizawa, A., Goto, S. G., & Nakashima, Y. (2019). "A nudibranch removes rival sperm with a disposable spiny penis." Journal of Ethology, 37(1):21-29. https://link.springer.com/article/10.1007/s10164-018-0562-z
[4] Lange, R., Werminghausen, J., & Anthes, N. (2014). "Cephalo-traumatic secretion transfer in a hermaphrodite sea slug." Proceedings of the Royal Society B: Biological Sciences, 281(1774):20132424. https://doi.org/10.1098/rspb.2013.2424
[5] Pennings, S. C. (1991). "Reproductive behavior of Aplysia californica Cooper: diel patterns, sexual roles and mating aggregations." Journal of Experimental Marine Biology and Ecology, 149:249-266.
[6] Leonard, J. L. (1990). "The Hermaphrodite's Dilemma." Journal of Theoretical Biology, 147(3):361-371.
[7] Bateson, G. (1936). Naven: A Survey of the Problems Suggested by a Composite Picture of the Culture of a New Guinea Tribe Drawn from Three Points of View. Cambridge University Press.
[8] アーヴィング・ゴッフマン著、丸木恵祐・本名信行訳『集まりの構造―新しい日常行動論を求めて』誠信書房、1980年
すでに登録済みの方は こちら









読者限定の内容も逃しません。